
Жевательная мускулатура хомякообразных Cricetidae.
Жевательная мускулатура млекопитающих представлена несколькими самостоятельными мускулами, каждый из которых в свою очередь имеет сложную внутреннюю дифференцировку. Взаиморасположение мускулов и их частей, степень их развития и направление волокон в конечном итоге отражают пищевую специализацию данного вида. Благодаря сложности строения жевательной мускулатуры и тесной взаимосвязи ее частей более или менее значительное изменение челюстного аппарата, связанное либо с переходом к новому типу питания, либо с приобретением дополнительных функций, ведет к преобразованию жевательной мускулатуры. В то же время в эволюции жевательной мускулатуры значительная роль принадлежит исторической преемственности структуры мускульной дифференцировки.
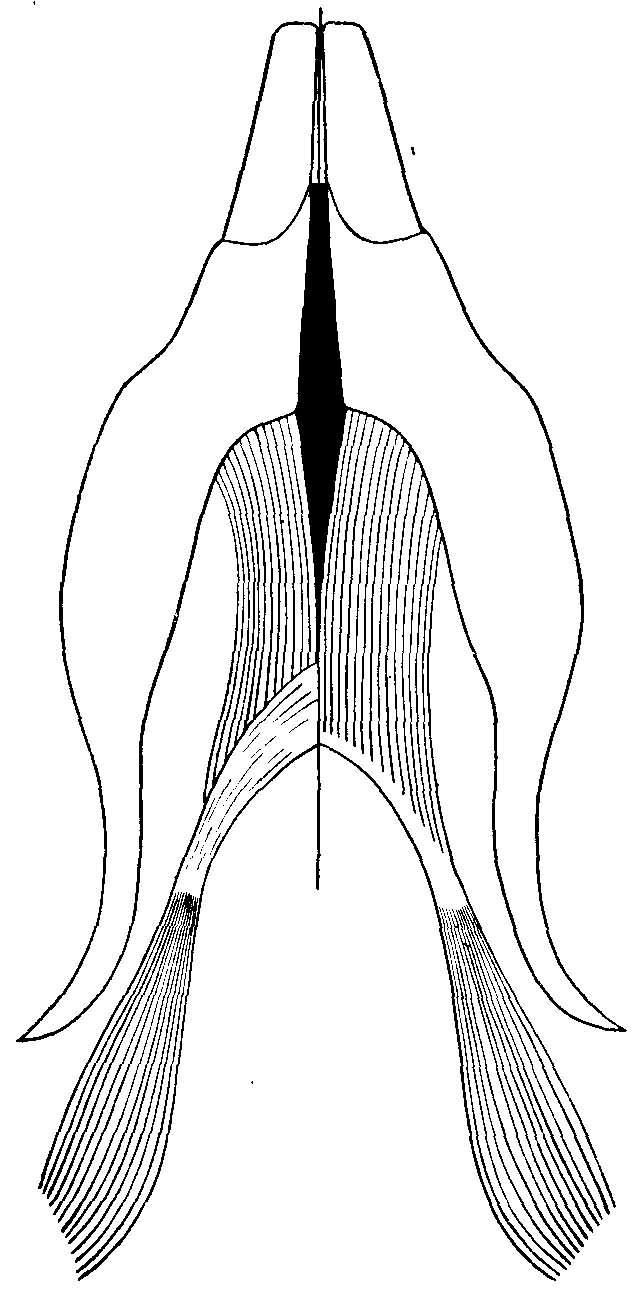
Рис. 1. Сравнение строения m. digastricus у Neotoma sp. — справа и Sigmodon hispidus Say et Ord — слева (из Rinker, 1954). Вид с вентральной стороны.
В процессе адаптивной радиации млекопитающих преобразование жевательной мускулатуры шло на фоне сохранения общего плана строения. Благодаря этому удается реконструировать последовательные стадии ее преобразования и проследить ее филогенетическое развитие. Это обстоятельство дало возможность разделить грызунов на крупные таксоны, исходя из особенностей конструкции их жевательной мускулатуры. Правомочность такого разделения вызывает вполне закономерный скепсис, так как вероятность независимого становления даже общего плана строения жевательной мускулатуры довольно велика. Тем не менее спорность объема выделенных таким путем крупных таксонов ничуть не умаляет значения изучения жевательной мускулатуры для филогенетических построений; это обстоятельство лишний раз подтверждает неправомерность построения системы на основании исследования развития лишь одной группы признаков, даже столь существенных, как строение жевательной мускулатуры.
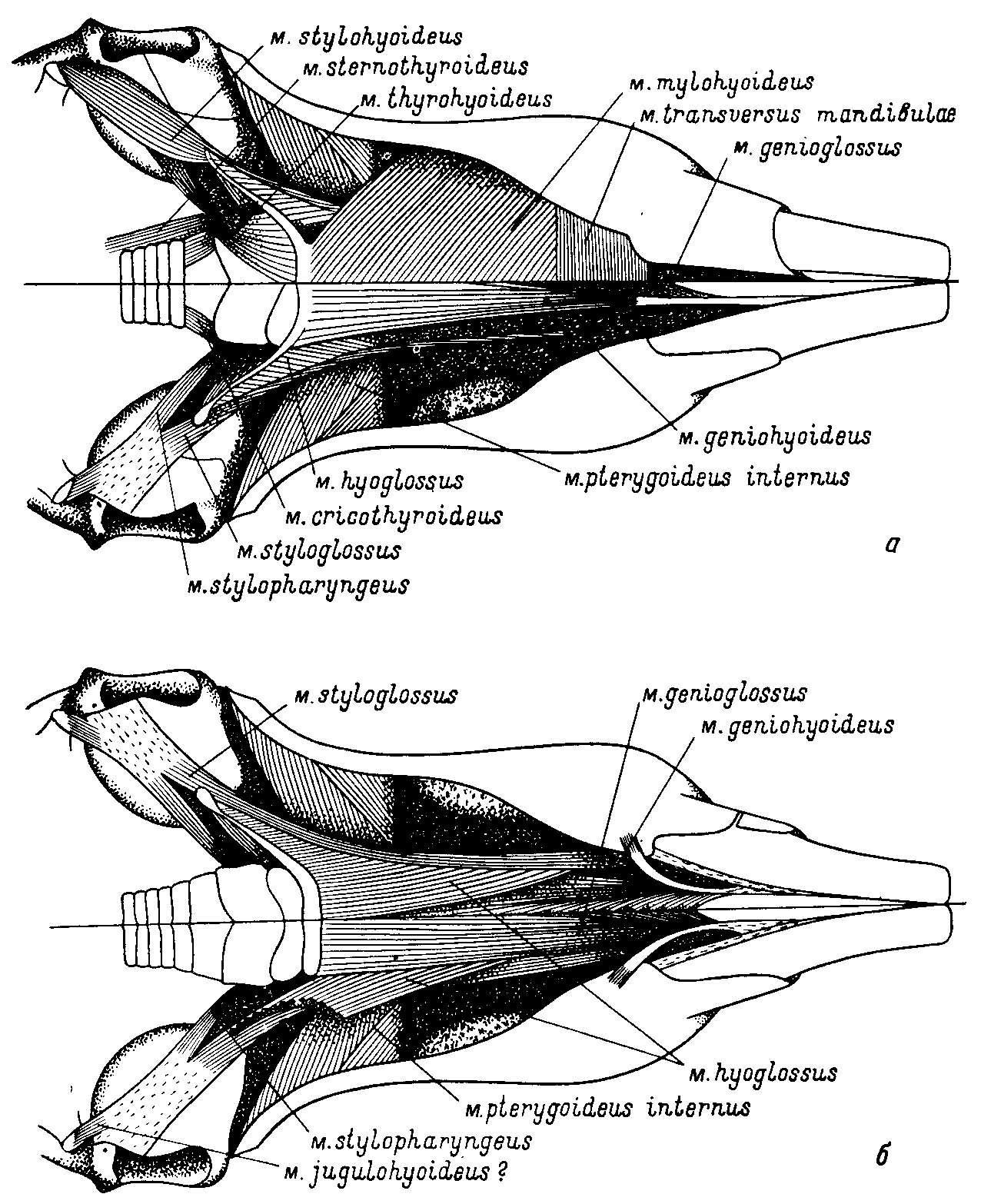
Рис. 2. Мускулатура нижней челюсти и подъязычного аппарата Sigmodon hispidus Say et Ord (из Rinker, 1954). Вид с вентральной стороны, а — верхняя половина рисунка — поверхностная мускулатура, нижняя — более глубокие части после удаления m. transversus mandibulae, m. mylobyoideus и m. stylohyoideus; б — верхняя половина рисунка — глубокие слои мускулатуры, нижняя — самые глубокие слои мускулатуры.
Учитывая то ключевое положение, которое занимает челюстной аппарат в целом и жевательная мускулатура в частности в комплексе адаптаций к питанию, и важность материала, который может быть получен при изучении этой системы, кажется поразительным столь сильное отставание сравнительной миологии черепа от сравнительной морфологии других органов.
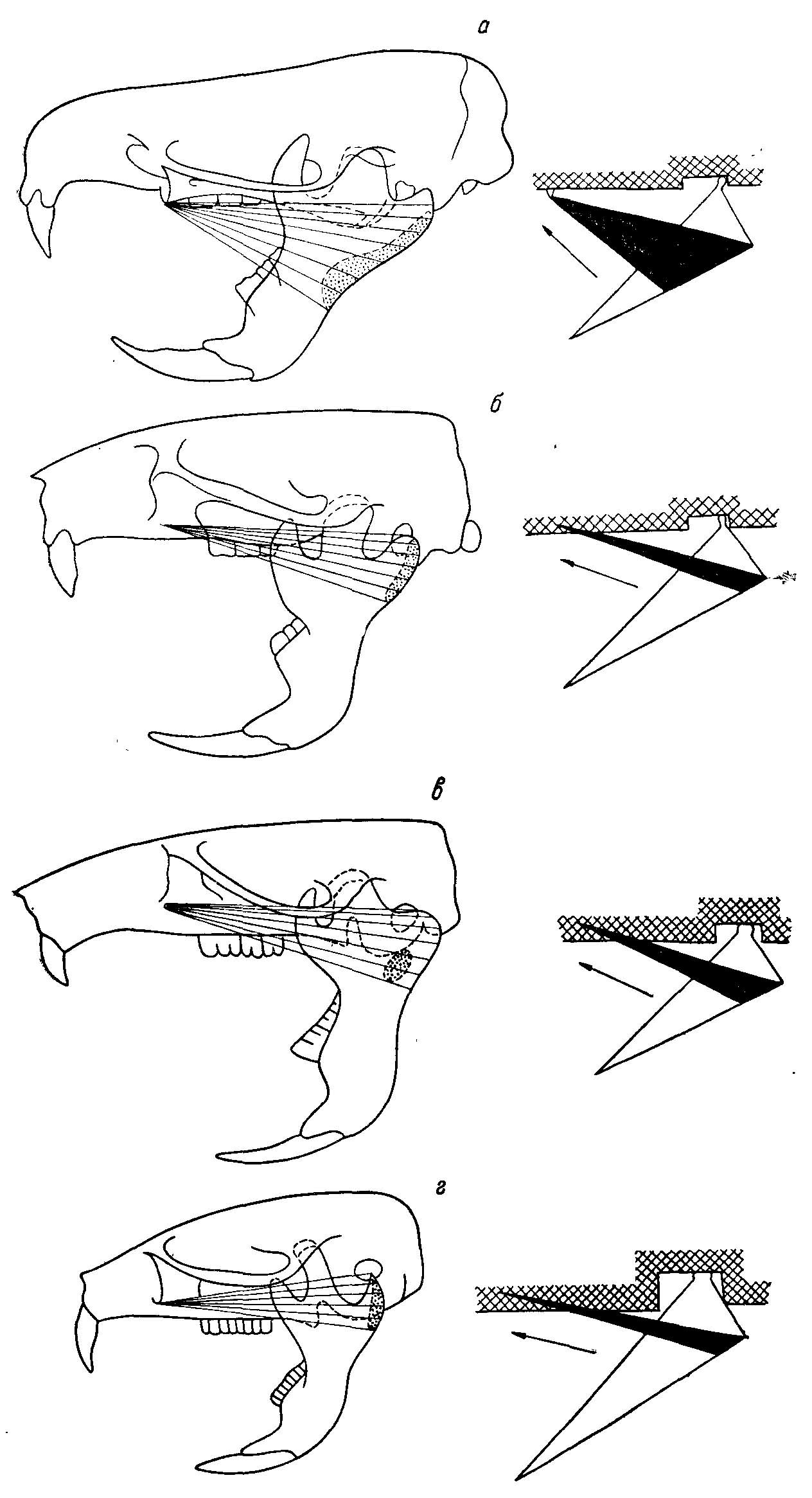
Рис. 3. Изменение функций m. mass. lateralis pars superficialis. Пунктиром показаны места прикрепления мышцы на внутренней стороне челюсти. Справа — схемы действия наружной поверхностной порции жевательной мышцы (у полевок эта мышца участвует только в продольноперетирающих движениях) (из Воронцова, 1963ж). а — Ichthyomys soderstrdmi de Winton; б — Cricetus cricetus L.; в — Neotoma cinerea Ord; a — Microtvs agrestis L.
Сведения по жевательной мускулатуре хомякообразных содержатся в ряде работ. Тулльберг (Tullberg, 1899) описал строение жевательной мускулатуры 6 видов хомякообразных, в том числе и 3 видов Cricetinae. Сравнительный анализ миологии 5 родов американских Cricetinae проведен Ринкером (Rinker, Hooper, 1950; Rinker, 1954, 1963). Описание жевательной мускулатуры Neotoma дано в работе Ховела (Howell, 1926). Корреляции между элементами черепа и жевательной мускулатурой на примере Ondatra zibethica рассмотрены Венделоо (Vendeloo, 1953). Жевательная мускулатура полевок изучена Репеннингом (Repenning, 1968), а слепышовых — Мегели и Крапом (Mehely, 1909; Кгарр, 1965). Анализ строения жевательной мускулатуры Mesocricetus в связи с особенностями питания проведен Горниаком (Gorniak, 1978). Анализ механизма жевательных движений, строения черепа и жевательной мускулатуры, некоторых закономерностей преобразования челюстного аппарата и черепа у грызунов и, в частности, Cricetidae дан в предыдущих исследованиях автора (Воронцов, 1961б, 1962а, 1962б, 1963ж, 1967а, 1967б; Воронцов, Лабас, 1968).
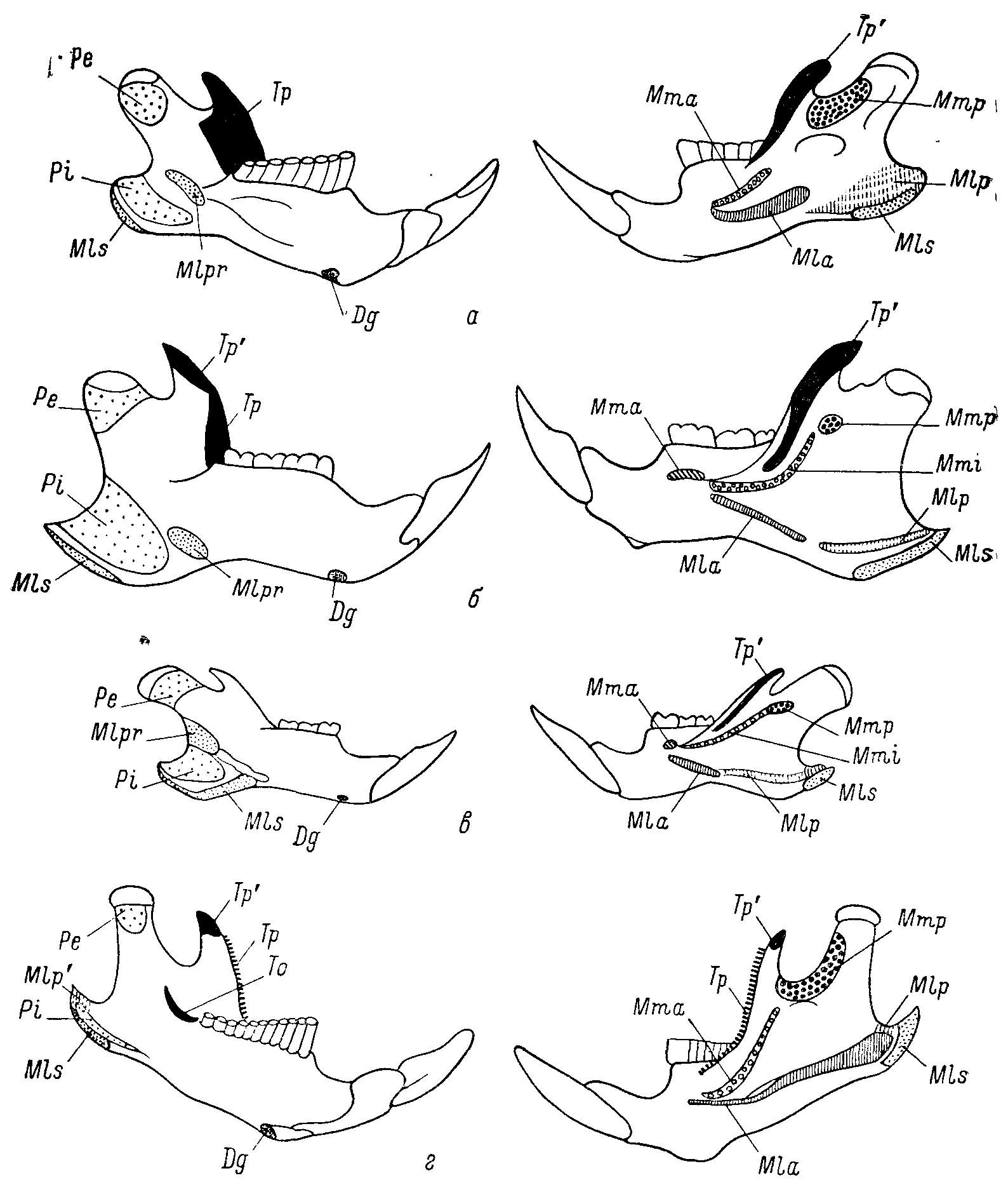
Рис. 4. Изменения мест крепления жевательных мускулов на нижней челюсти у Cricetinae, Nesomyinae и Microtinae. а, г — по Repenning, 1968; б, в — ориг. Левые половины нижних челюстей: левый ряд — вид с медиальной стороны, правый ряд — вид с латеральной стороны, а — Cricetinae — Neotoma mexicana Baird; б, в — Nesomyinae: б — Brachyuromys betsileoensis Bartl., в — Macrotarsomys bastardi Milner Edw. et Grandid.; г — Microtinae — Microtus longicaudus Merriam. Обозначения мест крепления мышц. Dg — m. digastricus, Mia — m. masseter lateralis profundus pars anterior, Mlp и Mlp' — разные области крепления m. mass. lateralis profindus pars posterior, Mis — m. mass. lat. pars superficialis, Mlpr — pars reflexa, Mma — m. mass. medialis pars anterior, Mmi — m. mass. medialis pars intermedius, Mmp — m. mass. medialis pars posterior, Pe — m. pterygoideus externus, Pi — m. pterygoideus internus, To — m. temporalis pars orbitalis, Tp и Tp' — разное области крепления m. temporalis pars profundus и pars superficialis.
Общие понятия, терминология.
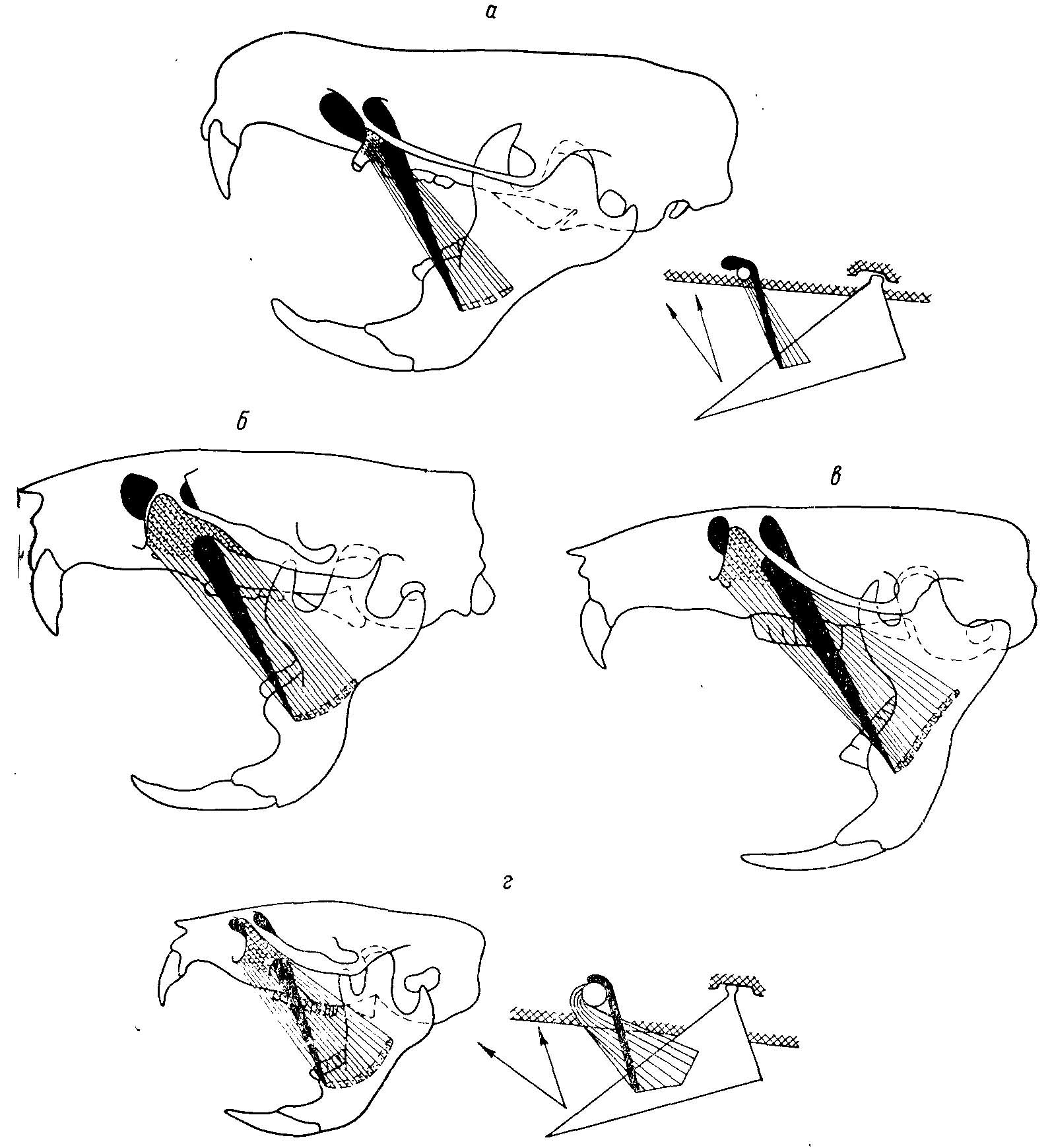
Рис. 5. Преобразование функций мышц, приводящих нижнюю челюсть в ряду Cricetidae (из Воронцова, 1963ж). а — Ichthyomys soderstromi de Winton; б — Cricetus cricetus L.; в — Neotoma cinerea Ord; г — Microtus agrestis L. Черным показан ход m. mass. medialis pars anterior; штриховкой — ход m. mass, lateralis profundus pars anterior; пунктиром — места крепления этой мышцы на черепе. Справа схемы действия мышц у крайних членов ряда. Видно уменьшение относительных размеров, перенесение начала m. mass. medialis pars anterior вверх и назад и перенесение места крепления m. mass, lateralis profundus pars anterior на черепе вперед и вверх, и на челюсти — назад и перпендикулярно вверх.
Челюстной аппарат обслуживается парными m. masseter, m. temporalis, m. pterygoideus, m. digastricus и непарным m. transversus.
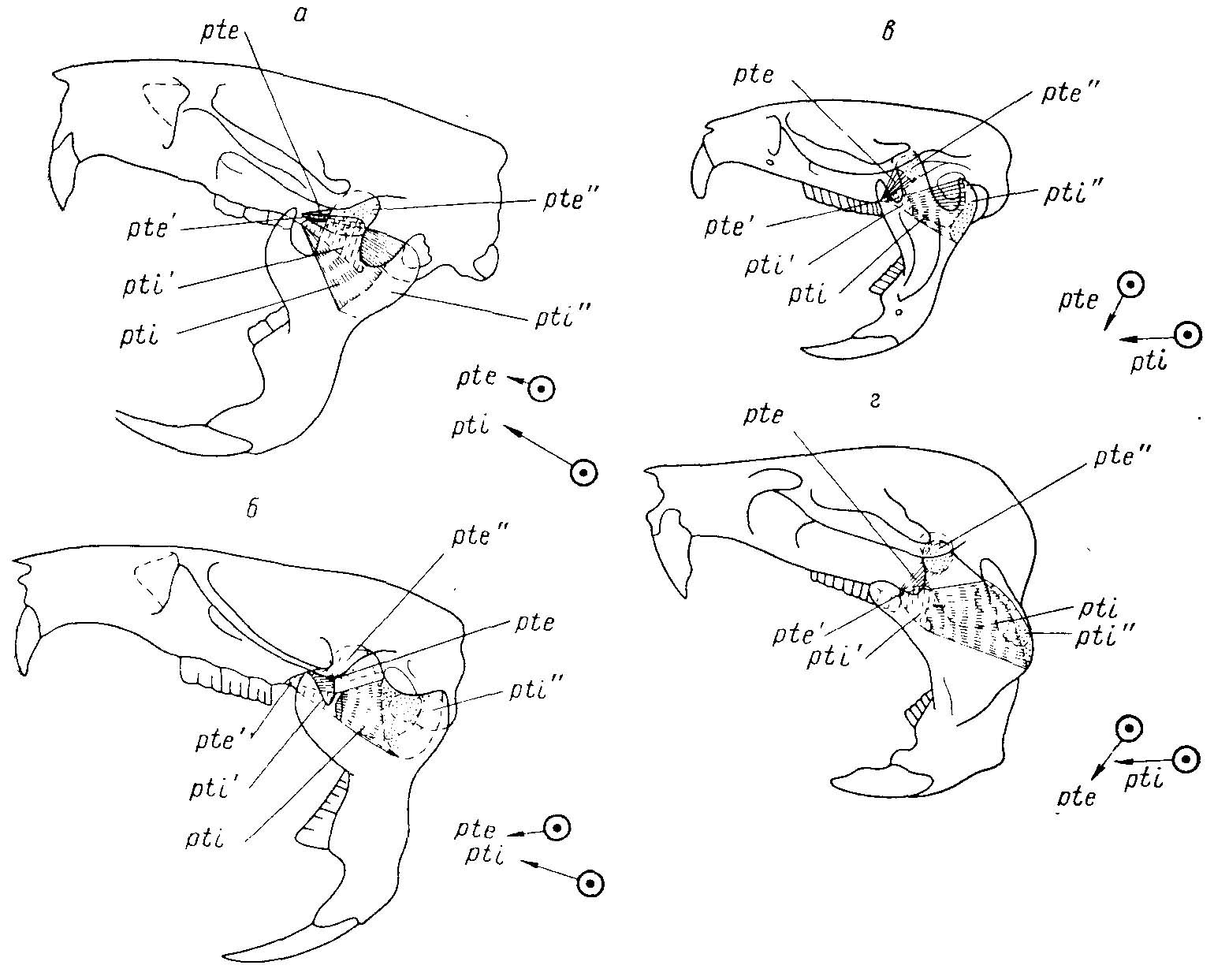
Рис. 6. Изменение мест прикрепления и функций крыловидных мышц в ряду Cricetidae (из Воронцова, 1963ж). а — Cricetus cricetus L.; б — Neotoma cinerea Ord; в — Microtus agrestis L.; г — Myospalax fontanier Milne-Edw. Пунктир — места прикрепления мышц на нижней челюсти; вертикальная прерывистая штриховка — места крепления мышц на черепе; стрелки — направление действия крыловидных мышц; pte, pte' и pte" наружная крыловидная мышца, ее крепление на черепе и челюсти; pti, pti' и pti" — внутренняя крыловидная мышца и площадки ее крепления на черепе и челюсти. Наружная крыловидная мышца, первично участвующая в поперечном смещении челюсти (а), затем выполняет функцию отведения задней части нижней челюсти вниз (в, г). Внутренняя крыловидная мышца, первично участвующая в приведении нижней челюсти и незначительной подаче ее вперед для грызения в переднем положении (а), затем принимает участие в подаче челюсти вперед (в, г).
Дериватами дорсальной группы констриктора I жаберной дуги у млекопитающих являются m. masseter, m. temporalis, m pterygoideus, иннервируемые жевательной ветвью тройничного нерва (ramus mandibulae n. trigeminus V). М. transversus и переднее брюшко m. digastricus, иннервируемые милогиоидной веточкой жевательной ветви тройничного нерва, — дериваты вентральной группы констриктора I жаберной дуги. Мускулатура II жаберной дуги в жевательной мускулатуре млекопитающих представлена лишь задним брюшком m. digastricus, которое иннервируется двубрюшной веточкой лицевого нерва n. facialis VII.
Если выделение в комплексе жевательной мускулатуры отдельных мышц не вызывает сомнений, то практически невозможно найти двух исследователей, придерживающихся одинаковых взглядов при идентификации и гомологизации порций и частей каждого мускула. Чтобы не увеличивать без того длинный список схем расчленения жевательных мышц на части, мы воспользовались номенклатурой и схемой дифференциации жевательной мускулатуры, предложенными Ринкером (Rinker, 1954).

Рис. 7. схема разделения жевательных мышц хомякообразных на части.
При описании мускулатуры крепление мускула к осевому черепу мы принимаем за начало, а крепление к нижней челюсти рассматриваем как окончание.
Принципиальные особенности строения жевательной мускулатуры едины для всех современных представителей надсемейства Muroidea. Однако при общности плана строения адаптивные изменения, связанные с пищевой специализацией, приводят к заметному разнообразию дифференцировки жевательных мускулов.
Принципиальная схема строения жевательной мускулатуры хомякообразных в целом сводится к следующему.
I. М. Digastricus (рис. 1). Двубрюшный мускул двойственного происхождения. Заднее брюшко начинается от proc. parooccipitalis (=jugularis) непосредственно и с помощью апоневроза aj (aponeurosis jugularis). Апоневроз, на котором оканчивается заднее и начинается переднее брюшко, крепится к поверхности basihyale. Заднее брюшко оканчивается на задней лопасти апоневроза adg (ар. digastricus) и его гребне, разделяющем переднее и заднее брюшки m. digastricus. В некоторых случаях оба брюшка ясно не обособлены — мускульные волокна переднего брюшка продолжаются в заднем.
Переднее брюшко начинается на передней стороне гребня апоневроза adg и тянется к вентральной поверхности нижней челюсти, где крепится позади симфиза в основном с помощью апоневроза am (ар. mandibularis).
Передние брюшки правой и левой стороны либо смыкаются, либо тянутся отдельно друг от друга.
М. digastricus отводит нижнюю челюсть и открывает рот.
II. М. transversus mandibulae (рис. 2) соединяет обе половинки нижней челюсти, крепится позади симфиза мандибул. Ход волокон параллельный. Поперечный мускул несколько сводит ветви челюстей и разводит нижние резцы.
III. M. masseter. Жевательный мускул расположен латеральнее других мускулов и представляет собой пласт мускульных волокон, идущих от скуловой дуги к боковой поверхности нижней челюсти. Он снабжен большим количеством апоневрозов, формирующих его сухожильный каркас и определяющих характер его внутренней дифференцировки. В составе жевательного мускула выделяют три в большей или меньшей степени обособленные порции, соответствующие трем последовательным слоям: наружную поверхностную — m. masseter lateralis pars superficialis, боковую — m. masseter lateralis profundus и внутреннюю — m. masseter medialis. Две последние в свою очередь подразделяются на подчиненные части.
1. М. masseter lateralis pars superficialis обособлена лучше прочих порций жевательного мускула. Она начинается сухожилием, переходящим в апоневроз as (ар. superficialis) от небольшой площадки верхнечелюстной кости сразу же под предглазничным отверстием (foramen infraorbitale). Эта площадка отлично видна у всех грызунов, вследствие чего легко можно восстановить ход pars superficialis у ископаемых форм. Форма сухожилия в поперечном сечении может быть различной: от округлой и почти точечной до имеющей вид широкой плоской ленты. Оканчивается эта порция на вентролатеральном крае углового отростка нижней челюсти и боковой поверхности апоневроза alp (ар. lateralis posterior), перекидывается через нижний край челюсти и тянется по ее медиальной стороне вдоль желобка, ограничивающего снизу альвеолярный бугор резца. Эта часть мускула соответствует pars reflexa, описанной у дикообразовых (Tullberg, 1899). Тулльберг рассматривал эту особенность строения поверхностной порции жевательного мускула как характеристику «трибы» Hystricognathi. Впоследствии pars reflexa была найдена не только у Hystricomorpha (Forster, 1928-1929; Woods, 1972), но и у Sciurus и Aplodontia (Hill, 1937), Dipodomyinae (Howell, 1932), Dipodoidea (Klingener, 1964), Cricetinae (Rinker, 1954).
M. mass. lateralis pars superficialis в своей каудальной части смыкается с поверхностной головкой m. Mass. lateralis profundus. По границе между ними проходит жевательная ветвь тройничного нерва. М. mass. lat. pars superficialis по сравнению с другими порциями этого мускула располагается по отношению к продольной оси нижней челюсти наиболее полого. В некоторых случаях она тянется почти параллельно оси mandibulae (рис. 3). Основная ее роль заключается в подаче нижней челюсти вперед, хотя она участвует также и в приведении нижней челюсти.
2. М. masseter lateralis profundus образует второй, считая снаружи, слой мышц. Его части слабо обособлены друг от друга, но все-таки отличаются по направлению волокон. Обычно в составе этой порции различают две части: переднюю — pars anterior и заднюю — pars posterior.
Передняя часть боковой порции жевательного мускула (m. mass. lat. profundus pars anterior) начинается от так называемой массетерной площадки верхнечелюстной кости и заходит на переднюю часть скуловой дуги. Положение массетерной площадки на ramus inferior предглазничного отверстия различно у разных видов. В креплении к скуловой дуге могут участвовать один или два начальных апоневроза — ар. superficialis anterior (asa) и ар. profunda anterior (ара). Начало этой части может быть смещено на боковую поверхность рострума над предглазничным отверстием.
На нижней челюсти эта часть в основном крепится с помощью апоневроза ala (ар. lateralis anterior) к так называемому массетерному гребню, идущему от основания М1 к угловому отростку (рис. 4). Помимо этого, небольшая часть мускульных волокон pars anterior оканчивается непосредственно на кости между гребнями нижней челюсти и на боковой поверхности конечного апоневроза ama (ар. medialis anterior) передней части внутренней порции жевательного мускула.
По сравнению с другими частями боковой порции жевательного мускула pars anterior проходит наиболее круто по отношению к продольной оси черепа, что свидетельствует об ее участии преимущественно в приведении нижней челюсти (рис. 5).
Задняя часть бокового жевательного мускула (m. mass. lat. profundus pars posterior) непосредственно примыкает сзади к передней части и по сути дела образует с ней единый мускульный пласт. Направление волокон из вертикального в передней части сменяется на наклонное — в задней. В составе задней части в свою очередь удается выделить 2 мускульные доли: поверхностную — m. mass. lat. profundus pars posterior superficialis и глубокую — m. mass. lat. profundus pars posterior profundus («superficial division» и «deep division», по Woods, 1972). Первая соответствует задней части поверхностной порции жевательной мышцы (m. mass. lat. superficialis pars posterior), в трактовке Мегели (Mehely, 1909). Эта головка представляет собой одноперистый мускул с апоневрозом asp (ар. superficialis posterior) в роли стержня, отсылающего волокна к боковой поверхности апоневроза alp и к самому концу углового отростка. Более наклонное положение волокон отличает ее от других частей боковой порции жевательного мускула. Она проходит почти так же полого, как и поверхностная порция, с которой она смыкается в каудальной части. Это дало основание Мегели (Mehely, 1909) отнести эту часть не к боковой, а к поверхностной порции в качестве ее задней части.
Благодаря наклонному положению поверхностной доли ее функция не ограничивается лишь приведением нижней челюсти, но в значительной степени включает подачу челюсти вперед.
Глубокая головка задней части (или pars posterior profundus) представлена двуперистым мускулом со стержневым апоневрозом арр (ар. profunda posterior). Волокна от боковой поверхности апоневроза арр оканчиваются главным образом на внутренней стороне апоневроза alp, тогда как от внутренней его поверхности мускульные волокна идут в вентро-каудальном направлении непосредственно к боковой поверхности нижней челюсти. Площадь, занимаемая задней частью на нижней челюсти, у разных видов различна. Помимо описанных выше апоневрозов, в сухожильный каркас задней части может включаться еще один апоневроз — ali (ар. lateralis insertionis).
В некоторых случаях в задней части боковой порции жевательного мускула условно можно выделить дополнительную лопасть, примыкающую каудально к pars posterior и проходящую более круто. Начинается она от скулового отростка squamosum и небольшого апоневроза арр’, являющегося продолжением апоневроза арр. Оканчивается непосредственно на дорсальном крае углового отростка или на апоневрозе ас (ар. caudalis).
В целом боковая порция проходит более круто по отношению к продольной оси черепа, чем поверхностная, и в основном участвует в приведении нижней челюсти.
3. М. masseter medialis — самый глубокий слой жевательного мускула. Эта порция представляет собой пласт мускульных волокон, веерообразно сходящихся к нижней челюсти. Направление волокон по мере продвижения назад изменяется от вентрокаудального к вентроростральному. Начало этой порции занимает всю внутреннюю поверхность скуловой дуги. В составе m. mass. medialis выделяют две части: переднюю — pars anterior и заднюю — pars posterior, которая в свою очередь в некоторых случаях подразделяется на две части: pars posterior собственно и промежуточную — pars intermedius.
В роли m. mass, medialis pars anterior выступает мускульный пучок идущий через предглазничное отверстие от боковой поверхности maxillare и intermaxillare, а также волокна, начинающиеся на внутренней поверхности ramus inferior скуловой дуги. К нижней челюсти pars anterior крепится с помощью апоневроза ama (ар. medialis anterior), впереди смыкающегося с апоневрозом ala.
Наибольшие затруднения возникают при разграничении передних частей боковой и внутренней порций жевательного мускула. Репеннинг (Repenning, 1968) относит к передней части бокового массетера (m. mass. lat. profundus pars anterior) волокна, собирающиеся на апоневрозе ala, оставляя за m. mass. medialis pars anterior часть мускульного пласта, идущего к апоневрозу ama. Граница в мускульной массе между ними условна.
М. mass. medialis pars anterior приводит и лишь едва подает вперед нижнюю челюсть (рис. 5).
Задняя часть внутренней порции (m. mass. medialis pars posterior) начинается от медиовентральной стороны jugale и скулового отростка (processus zygomaticus) чешуйчатой кости. Оканчивается она на боковой поверхности венечного отростка и в ямке, расположенной над альвеолярным резцовым бугром. Pars posterior может быть снабжена двумя апоневрозами — (amo) (ар. medialis originalis) и (amp) (ар. medialis posterior).
Часть мускульных волокон от середины скуловой дуги, идущая к наружной поверхности основания венечного отростка, может выделяться как pars intermedius (Rinker, 1954).
Pars posterior приводит и несколько сдвигает назад нижнюю челюсть.
IV. М. temporalis. Если жевательный мускул приурочен в основном к латеральной поверхности нижней челюсти, то височный мускул (m. temporalis) топографически связан с ее медиальной стороной. В отличие от жевательного, височный мускул более компактен и может быть подразделен на части весьма условно.
М. temporalis представлен мускульным пером, волокна которого веерообразно сходятся на конечном апоневрозе at (ар. temporalis). Начинается этот мускул на parietale, interparietale, squamosum, может покрывать заднюю часть frontale. Теменные гребни довольно точно обрисовывают верхнюю границу крепления этого мускула. Несмотря на внешнюю монолитность височного мускула, в его составе выделяют две, а некоторые авторы даже три части.
Существуют несколько подходов к выделению частей этого мускула. Ринкер (Rinker, 1954) различает в составе m. temporalis переднюю поверхностную часть, которую мы будем называть pars superficialis, и заднюю, более глубокую часть — pars profundus.
Мускульные волокна поверхностной части (m. temporalis pars superficialis) тянутся от верхнего края теменного гребня и от основания апоневроза ats (ар. temporalis superficialis) к боковой поверхности апоневроза at и к переднему краю венечного отростка.
Основная масса височного мускула принадлежит его глубокой части (m. temporalis pars profundus). Она начинается от теменного и ламбдоидного гребней, в передней части подстилает pars superficialis и заполняет всю височную впадину. Мускульные волокна pars profundus собираются на медиальной поверхности нижней челюсти в ямке, находящейся между основанием венечного отростка и at. В креплении глубокой части к осевому черепу могут участвовать апоневрозы atm (ар. temporalis medialis), идущий от ламбдоидного гребня, и atp (ар. temporalis profundus), подстилающий мускул изнутри и начинающийся на задней стенке глазницы.
Конечный апоневроз at крепится вдоль переднего края венечного отростка. У основания proc. coronoideus апоневроз at загибается на медиальную поверхность мускула и оканчивается на внутренней стороне нижней челюсти, принимая волокна от стенки мозговой капсулы и апоневроза atp. Увеличенная лопасть апоневроза at, которая в некоторых случаях обособляется в самостоятельный апоневроз atc (ар. temporalis caudalis), собирает на свою медиальную поверхность волокна, идущие почти горизонтально от ламбдоидного и задней части теменного гребней. Это мускульное полуперо иногда противопоставляется всему остальному височному мускулу и трактуется как pars posterior m. temporalis, тогда как передней части pars profundus совместно с pars superficialis (в трактовке Ринкера (Rinker, 1954)) присваивается статус передней части височного мускула — pars anterior.
У некоторых хомякообразных происходит обособление еще одной части, которую мы называем pars orbitalis. Она тянется от задней стенки глазницы и глубокого апоневроза atp и оканчивается обособленно от pars profundus у основания последнего коренного зуба. Эта часть снабжена самостоятельным конечным апоневрозом ato (ар. temporalis orbitalis), обособившимся от внутренней части апоневроза at.
М. temporalis приводит и несколько сдвигает назад нижнюю челюсть.
V. М. pterygoideus. В отличие от жевательного и височного мускулов, крыловидный мускул (m. pterygoideus) относится к группе вентральных аддукторов. По сути дела m. pterygoideus состоит из двух самостоятельных мускулов, выполняющих различные функции: наружного — m. pterygoideus externus и внутреннего — m. pterygoideus internus.
1. M. pterygoideus externus начинается на боковой поверхности alisphenoideum и боковой поверхности крыловидного отростка задней части palatinum и заполняет промежуток между bulla tympani и задним краем М3. Начинаясь плоской и сравнительно широкой лентой на черепе, m. pterygoideus externus затем сужается и приобретает округлую в сечении форму.
На нижней челюсти этот мускул крепится на медиальной стороне ргос. condyloideus под сочленовной головкой. В креплении к осевому черепу и к нижней челюсти могут участвовать апоневрозы аео (ар. externa originalis) и aei (ар. externa insertionis).
М. pterygoideus externus в основном участвует в поперечных движениях нижней челюсти.
2.М. pterygoideus internus (внутренний крыловидный мускул) отделен от наружного. Он начинается от боковой крыловидной ямки (fossa pterygoidea lateralis) и крепится на широкой площадке с медиальной стороны углового отростка, ограниченной сверху альвеолярным выступом нижнего резца, а снизу — загнутым внутрь нижним краем углового отростка. Внутренний крыловидный мускул претерпевает значительные измёнения в ряду хомякообразных (рис. 6).
В составе m. pterygoideus internus можно выделить две части: вентральную и дорсальную. Вентральная часть имеет сложную внутреннюю дифференцировку и объединяет несколько перистых структур, направленных под некоторым углом навстречу друг другу. Ее сухожильный каркас представлен двумя конечными апоневрозами: поверхностным вентральным avs (ар. ventralis superficialis) и глубоким вентральным avp (ар. ventralis profundus), которые крепятся к медиальному краю углового отростка нижней челюсти, и начальным апоневрозом avo (ар. ventralis originalis). Апоневроз avo вклинивается между конечными апоневрозами. Мускульные волокна, идущие от апоневроза ado к дорсальной поверхности апоневроза avp, как правило, отличающиеся по направлению от остальной части pars ventralis, можно обозначить как m. pter. internus pars intermedius.
Дорсальная часть m. pterygoideus internus начинается апоневрозом ado (ap. dorsalis originalis) от боковой крыловидной пластинки и отсылает волокна ко внутренней стороне углового отростка, к которому крепится либо непосредственно, либо с помощью апоневроза adi (ар. dorsalis insertionis).
Внутренние крыловидные мускулы, сокращаясь одновременно, приводят нижнюю челюсть и немного подают вперед. При поочередном сокращении m. pterygoideus internus участвует в поперечных перетирающихся движениях.
Таким образом, схема разделения жевательных мышц на части, принятая в данной работе, сводится к рис. 7.
Источник: Н. Н. Воронцов. "Фауна СССР. Млекопитающие. Том III, вып. 6. Низшие хомякообразные (Cricetidae) мировой фауны", 1982 г.
Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
Зверосайт в соцсетях:



Зверо-Видосы:





"вы теряете клиентов продвинь свой сайт в топ один за десять минут тг @Spd_Top" пишет на странице: Муфлон.
19.06.2026 11:49:28
вы теряете клиентов продвинь свой сайт в топ один за десять минут тг @Spd_Top
"продвинь свой сайт в топ один за десять минут тг https://t.me/Spd_Top" пишет на странице: Муфлон.
12.06.2026 22:08:37
продвинь свой сайт в топ один за десять минут тг https://t.me/Spd_Top
"admin" пишет на странице: Лягушки веслоноги (Theloderma) и их разведение.
31.03.2026 06:47:46
Нет, не разводим, просто статья из журнала
"zukovskaarimma7@gmail.com" пишет на странице: Лягушки веслоноги (Theloderma) и их разведение.
19.03.2026 10:25:49
Вы разводите веслонога, можно приобрести?
"Маина" пишет на странице: Сколопендра гигантская.
09.03.2026 15:33:13
мечтаю о сколопендре. но мечтаю зря, поскольку дома и кота завести нельзя
Последние 11 статей:
- Амурский тигр выслеживает пятнистых оленей.
- Моржонок с мамой плавают в открытом океане.
- Моржи ютятся на тающих льдинах в поисках еды.
- Дикие кабанята сосут молоко у мамы.
- Как спасли очковых медведей.
- Стадо кабанов с детишками гуляют и ищут еду.
- Тюлень попался в пасть белой акулы?
- Пиявки умеют прыгать. Акробатика этих существ вызывает споры уже более ста лет.
- Белая акула сожрала котика за один укус.
- Пятнистые олени в лесу
- Осень на Дальнем Востоке России.














.")
.")
