
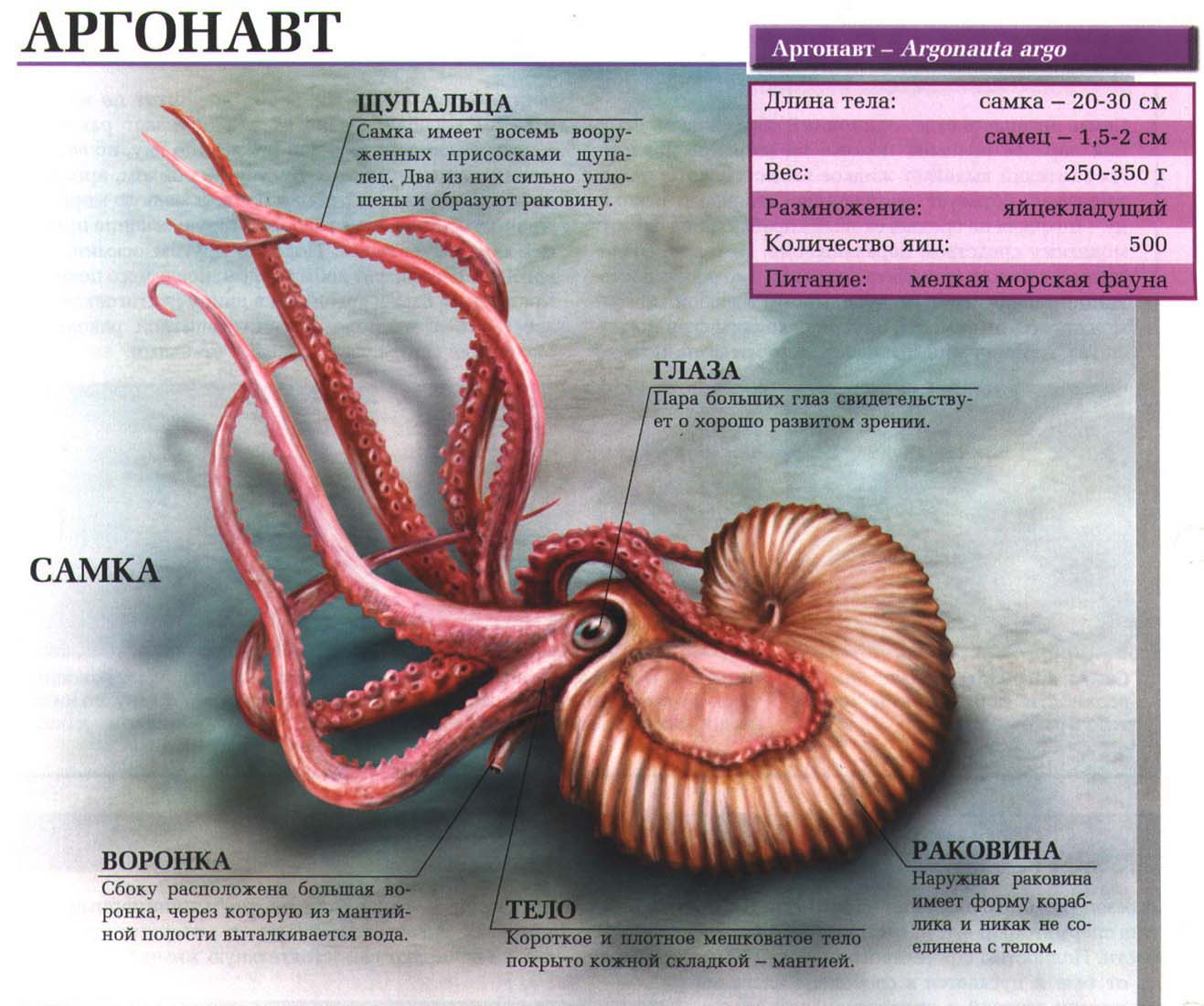
Первые млекопитающие. Как они появились и развивались?
История эволюции — история успеха. Жизнь приспособилась к самым сложным условиям, образуя великое видовое разнообразие. Узнавая о количестве и отличиях жизненных форм, можно глубже и лучше осознать величие эволюционного процесса. Сведения о них помогают понять, как происходит физическая адаптация и как существа приобретают тот или иной облик и те качества, которыми они обладают в наши дни.

Первые млекопитающие.
Среди самых успешных и известных вымерших животных были динозавры, правившие планетой на протяжении 150 миллионов лет. На настоящий момент их известно более 600 видов. Однако тех, кто в мезозое лишь ловко шнырял среди когтистых лап динозавров и кто впоследствии занял место «ужасных ящериц», вопреки расхожему мнению, было несравненно больше: от былого величия динозавров остались лишь груды костей, а природа шагнула дальше, вручив скипетр владычества млекопитающим.

Репеномам (Repenomamus).
В эволюционном учении часто употребляют термины «древние», «ископаемые», «примитивные», хотя это не совсем правильно. Современные мечехвосты не менее примитивны, чем ископаемые, первые насекомые не уступят по сложности нынешним формам, а третичные летучие мыши уже обладали сформированным летательным аппаратом и системой эхолокации. Латимерия, гинкго, лингула и наутилус существуют до сих пор. В последнее время даже орангутана причисляют к «живым ископаемым», обнаружив у него много общих признаков с жившим 14 миллионов лет назад рамапитеком. Это абсолютно справедливо и в отношении мезозойских млекопитающих, имевших в те времена не менее богатое морфологическое разнообразие, чем доминировавшие в воде, воздухе и на суше рептилии.

Реконструкция головы циногната (Cynognathus), который является представителем подотряда цинодонтов.
Предполагается, что млекопитающие произошли от синапсидных рептилий. Процесс маммализации, скорее всего, занимал почти весь триасовый период, т.е. около 40 миллионов лет, и осуществлялся параллельно у нескольких групп синапсид. Многие эволюционные ветви, даже накопив много прогрессивных признаков, в итоге оказывались тупиковыми, однако некоторым в конце триаса удалось выйти на новый эволюционный виток, в итоге превратившись в настоящих млекопитающих. Важнейшими маммальными признаками считаются:
Заламбдалест (Zalambdalestes).
- верхние обонятельные раковины;
- снабженные мускулатурой и чувствительными вибриссами губы;
- трехбугорчатые щечные зубы;
- челюстное сочленение зубной и чешуйчатой костей;
- слуховая система, состоящая из трех костей;
- крупные полушария телэнцефалического мозга.

Реконструкция морганукодона и среды его обитания.

Морганукодон (Morganucodon).

Фруитафоссор (Fruitafossor).

Тениолаб (Taeniolabis).
Касторкауда (Castorcauda).

Хальданодон (Haldanodon).
Самый первый признак наличествовал уже у раннепермских териодонтов (Theriodontia).
Птилод (Ptilodus).
Второй присутствовал у более поздних тероцефалов (Therocephalia). Циногнаты (Cynodontia, Cynognathidae) триасового периода обладали уже почти всеми признаками, отличаясь от ранних млекопитающих лишь строением челюсти и слухового аппарата. А вот формирование телэнцефалического мозга, обладающего ассоциативными зонами, продолжалось весь мезозой.
Параллельно с вышеуказанным набором признаков прогрессировали и другие физиологические особенности — теплоизолирующий шерстный покров, функциональность млечных желез, расположение конечностей непосредственно под туловищем, возможность измельчения и пережевывания пищи. Самым известным родом мезозойских млекопитающих, появившимся в палеонтологической летописи около 210 миллионов лет назад и наиболее хорошо изученным, является морганукодон (Morganucodon), обнаруженный в 1949 году в раннеюрских слоях Уэльса. Впоследствии различные фрагменты остатков морганукодона и его родственников были найдены в позднем триасе Франции и Швейцарии, юре Великобритании, Китая, Индии, Африки и США, позволивших детально изучить большую часть его скелета.
Волатикотерий (Volaticotherium).
Череп морганукодона относительно тела выглядит крупным и длиннее и тоньше по сравнению с черепами цинодонтов (Cynodontia). Хотя конец морды известен плохо, можно полагать, что наружные ноздри сливались в одно отверстие. Предчелюстные и верхнечелюстные кости участвовали в образовании вторичного неба, над которым находилась обширная носовая полость — скорее всего, она была выстлана обонятельным эпителием. Слуховая капсула морганукодона, как и у всех млекопитающих, уже окостеневает единым образованием — околоушной костью. Крошечные ямки на морде говорят о наличии вибрисс, крупные глазницы — о сумеречной или ночной активности.
Морганукодон обладал двойным рептилийным челюстным сочленением между сочленовной и квадратной костями, еще не преобразованными в слуховые косточки. Челюстной сустав у примитивных млекопитающих располагался непосредственно перед затылочным мыщелком, но у продвинутых форм был смещен вперед. Его примитивное заднее положение, возможно, объясняется унаследованной от цинодонтов тесной связью между нижнечелюстными костями и слуховой капсулой. Нижняя челюсть морганукодона выглядела вполне типично для млекопитающих, однако сохраняла ряд сильно редуцированных костей, присутствовавших у цинодонтов.
Зубы разделялись на резцы, клыки, предкоренные и коренные. Характер их износа очень специфичен. Коронки довольно острые, они могли использоваться для прокусывания добычи, а зубная система функционально находилась на стадии эволюции прикуса. Геометрия износа показывает, что движение челюстей у морганукодонтид, по всей вероятности, напоминало известное у некоторых примитивных современных млекопитающих — к примеру, опоссума. Ветви челюсти соединялись непрочно, что позволяло им двигаться относительно независимо, и одной из самых характерных черт являлось то, что прикус осуществлялся в каждый момент только одной ветвью челюсти. Детские молочные зубы сменялись набором постоянных.
Грудные и поясничные позвонки у морганукодона отчетливо различались. Как и у позднейших млекопитающих, шейные позвонки характеризовались широким невральным каналом, что, вероятно, связано с развитием плечевого нервного сплетения. Наличествовал длинный и тонкий хвост. Конечности напоминали таковые у продвинутых цинодонтов, однако ряд признаков характерен исключительно для млекопитающих. Конфигурация суставных поверхностей указывает, что постановка и характер движения конечностей напоминали свойственные ныне живущим млекопитающим типа сумчатого опоссума или плацентарной тупайи. Первый, или большой, палец задней конечности, видимо, несколько противопоставлялся остальным. Этот признак, а также наличие хорошо развитых когтей указывают на возможный древесный образ жизни. Однако данные признаки могли также возникнуть в любой сложной для локомоции среде, заставляющей часто хвататься, цепляться и карабкаться, когда функциональные различия между наземным и древесным образами жизни невелики.
Итак, морганукодона можно представить как зверька 15—20 см длиной и весом около 20-30 г с узкой мордочкой и удлиненными челюстями, усаженными мелкими и острыми зубами, в общих чертах похожего на землеройку или мышь. Строение зубов показывает, что морганукодон был больше плотоядным, чем всеядным. За исключением некоторой специализации зубной системы, у морганукодона почти идеальный тип строения, предполагаемый для предка всех более поздних млекопитающих. Одна из характернейших черт ранних млекопитающих — довольно мелкие размеры. Такой размер облегчал преследование крошечной добычи, как правило, различных беспозвоночных, и позволял укрываться от нападения крупных хищников.
По пропорциям тела, размерам и зубному аппарату ранние млекопитающие напоминают современных насекомоядных. Предполагается определенная аналогия с ними как по образу жизни, так и по физиологии, включая метаболизм и температуру. По мнению некоторых исследователей, температура тела ранних млекопитающих составляла всего лишь около 30 °С. В любом случае, если эти животные поддерживали относительно постоянную температуру, им требовалась достаточно высокая интенсивность метаболизма. В связи с этим можно предположить у них наличие густого шерстного покрова. К сожалению, шерсть в ископаемом состоянии не сохраняется, и никаких признаков, указывающих на ее существование, не обнаружено. Волоски могли быть заметны на отпечатках лап, но таких мезозойских находок пока не описано.
Поддержание высокой температуры и эволюция молочных желез стали предпосылкой для развития примитивного типа размножения. Вероятно, раннемезозойские формы размножались подобно современным однопроходным, вылупляясь из яиц на очень ранней стадии развития. Как отмечал Хопсон (1973), незрелость древнейших млекопитающих при рождении можно также объяснить мелкими размерами взрослых особей и физиологическими ограничениями, возникающими вследствие высокой метаболической интенсивности — около 90% энергии пищи, потребляемой мелкими зверьками, используется на поддержание высокой температуры. Для их детенышей было физиологически невозможно поддерживать высокую температуру тела за счет собственной метаболической активности, ведь и детеныши современных землероек и мелких грызунов не эндотермны, а эктотермны. В поддержании высокой температуры тела они полностью зависят от родителей, и голая кожа позволяет им поглощать тепло более эффективно. Когда детеныши избавлены от этой необходимости, почти все съеденное используется для роста и развития. Став взрослыми, такие мелкие формы вынуждены питаться почти непрерывно.
Кроме всего прочего, мелкие размеры, требующие метаболической интенсивности, компенсирующей быструю потерю тепла, подразумевали и высокую эффективность пищевого поведения, основанного на лучшем восприятии информации и двигательной функции. Совершенствование обоняния, слуха, зрения и осязания приводило к дальнейшему развитию высших мозговых центров. Размеры мозговой коробки морганукодона крупнее, чем у его предков аналогичного размера, и это изменение приближало размеры его мозга к таковым у примитивных кайнозойских млекопитающих.
Наряду с типовой формой в семейство морганукодонтид входили такие представители, как эозостродон (Eozostrodon) из позднего триаса Великобритании, эритротерий (Erythrotherium) из ранней юры Южной Африки, обладающий самыми крупными коренными зубами из всех морганукодонтов хельветиодон (Helvetiodon) из позднего триаса Швейцарии, а также найденные в Индии гондванадон (Gondwanadon) и индотерий (Indotherium). Все они отличаются друг от друга некоторыми изменениями в морфологии зубов и нижнелюстного сочленения, которые впоследствии привели к появлению новых, более прогрессивных групп мезозойских млекопитающих.
В течение «века динозавров» млекопитающие внешне оставались малозаметными членами мезозойских биоценозов. Однако, при скрытном образе жизни, в сумеречные часы, среди них происходила интенсивная радиация. Именно тогда и произошло расхождение эволюционных ветвей, из которых впоследствии выделились однопроходные, сумчатые и плацентарные. Первые, отличающиеся рептильным строением половых путей,— плохо известны в эволюционной истории. В палеогеновых отложениях Южной Америки найден утконосый обдуродон (Obdurodon), в Австралии — его дальние родственники: меловой стероподон (Steropodon) и палеогеновый тейнолоф (Teinolophos). В миоценовой Австралии обитала крупная доисторическая ехидна мегалибгвилия (Megalibgwilia). Однако на основе примитивных признаков строения предполагается, что клоачные отделились гораздо раньше — в триасовом периоде.
Мезозойские зверьки имели множество боковых ветвей, которые вымерли, не оставив потомков. Одними из самых распространенных теплокровных животных мезозоя являлись триконодонты (Triconodonta), поднявшие вымпел истинных млекопитающих: они были небольшого размера, явные хищники, имевшие архаичный головной мозг и отличающееся строение коренных зубов. Самый крупный триконодонт — репеномам (Repenomamus), найденный в раннемеловых слоях Китая, достигал метровой длины и мог весить 14 кг, представляя реальную угрозу для молодых особей динозавров. Представители другой группы, докодонтов (Docodonta), были более специализированны. Для хальданодона (Haldanodon) из юра Западной Европы предполагался норно-роющий образ жизни, а строение касторкауды (Castorcauda) свидетельствовало, что это животное было хорошо приспособлено для плавания, напоминая современную выдру.
Представители аллотериев (Allotheria), или многобугорчатых (Multituberculata), тоже отличаются своеобразным строением зубов. Во многих отношениях аллотерии являются уникальными — варьирующие в размерах от мыши до сурка, это, фактически, первая группа растительноядных животных, достигшая доминирования. По своей физиологии они могли быть близки к современным, а их эволюционный успех на мелпалеогеновой границе, по-видимому, демонстрировал несомненную адаптацию к питанию покрытосеменными растениями. Характеризующийся древесным образом жизни крысоподобный птилод (Ptilodus), землеройный ламбдопсалис (Lambdopsalis), напоминающий бобра тениолаб (Taeniolabis) — все эти аллотерии просуществовали с юрского периода до эоцена и бесследно исчезли.
Заслуживают внимания и другие древние отряды. Это — прежде всего, юрские шуотерии (Shuotheridia), внешне и по образу жизни схожие с землеройками, у которых в отличие от современных млекопитающих режущая грань коренных зубов располагалась за трущей поверхностью (псевдотрибосфенический тип). Другая группа — симметродонты (Symmetrodonta), названные так по симметричной форме верхних и нижних коренных зубов. Наиболее эволюционно продвинутые представители симметродонтов — такие, как юрский кюнеотерий (Kuehneotherium) или меловой спалакотерий (Spalacotherium),— филогенетически уже были довольно близки к ныне живущим млекопитающим. А похожий на крысу перамус (Peramus), типовой представитель отряда Регатига. имевший конвергентное сходство с бандикутами, наиболее близок предкам сумчатых и плацентарных.
Поскольку палеонтологическая летопись ранних млекопитающих состоит большей частью из зубов и челюстей, зачастую трудно определить таксономическое положение некоторых групп или родов. Неизвестно систематическое положение планирующего подобно летяге волатикотерия (Volaticotherium) или юрского мирмекофага — фруитафоссоpa (Fruitafossor), так как многие из этих особенностей либо малопригодна для установления филогенетических связей, либо уникальны.
Как упомянуто выше, исходная дивергенция млекопитающих началась с начала мезозоя. На сегодняшний день известно более двух десятков линий, из которых только две — монотрематы и метатерии-эвтерии, считающиеся сестринскими, привели к сегодняшнему разнообразию. Уже в палеогене известно около 30 групп, достаточно четко различающихся на уровне семейств. Согласно последним данным, начало грызунам, зайцеобразным и прыгунчиковым дали заламбдалестиды (Zalambdalestidae) из позднего мела Монголии. Потомки сборной группы кондилартр (Condylarthra) впоследствии дивергировали на копытных, хоботных, трубкозубых и китообразных. Хищные, насекомоядные, рукокрылые и приматы разделились в ходе ранней радиации общего ствола, у корней которого стоял кенналест (Kennalestes). А происхождение неполнозубых и ящеров до сих пор остается загадкой — возможно, их предки были первыми обособившимися группами плацентарных млекопитающих.
Автор – Игорь Казанов. Рисунки Д. Богданова. Коллажи В. Евсеева. Источник: журнал «В мире животных» 2011, № 12.
Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
Зверосайт в соцсетях:



Зверо-Видосы:





"admin" пишет на странице: Лягушки веслоноги (Theloderma) и их разведение.
31.03.2026 06:47:46
Нет, не разводим, просто статья из журнала
"zukovskaarimma7@gmail.com" пишет на странице: Лягушки веслоноги (Theloderma) и их разведение.
19.03.2026 10:25:49
Вы разводите веслонога, можно приобрести?
"Маина" пишет на странице: Сколопендра гигантская.
09.03.2026 15:33:13
мечтаю о сколопендре. но мечтаю зря, поскольку дома и кота завести нельзя
"Полли" пишет на странице: Паук сенокосец.
03.09.2025 20:03:41
Интересно, очень и они бегают. Я видела как 2 самца дрались за самку. Победитель спариался. И потом долго не отходил от самки и других не подпускал
"Я верю в то,что они живы" пишет на странице: Стеллерова корова командорских островов.
30.03.2025 02:07:53
Последние 11 статей:
- Амурский тигр выслеживает пятнистых оленей.
- Моржонок с мамой плавают в открытом океане.
- Моржи ютятся на тающих льдинах в поисках еды.
- Дикие кабанята сосут молоко у мамы.
- Как спасли очковых медведей.
- Стадо кабанов с детишками гуляют и ищут еду.
- Тюлень попался в пасть белой акулы?
- Пиявки умеют прыгать. Акробатика этих существ вызывает споры уже более ста лет.
- Белая акула сожрала котика за один укус.
- Пятнистые олени в лесу
- Осень на Дальнем Востоке России.





.")










.")
.")
